Reproduced with the permission
of the Scottish Naturalist
Copyright:
May be used for private research. All other rights
reserved
By DAVID S. MARTIN
Loch Ness and Morar Project
ADRIAN
J. SHINE
Loch Ness and Morar Project
ANNIE DUNCAN
Hydroacoustic Unit, Department
of Biology,
Royal Holloway University of London
Introduction
The importance and biological
interest of the profundal faunas of Loch Ness (230
m) and Loch Morar (310 m) is that they inhabit depths
greater than 200 m or 300 m, which are deep on a
world scale. The
profundal sediments are also very extensive, occupying
over 50% of the area of Loch Ness greater than 150
m deep (Smith, Cuttle and Maitland, 1981) and 27%
greater than 200 m deep; the corresponding percentages
for Loch Morar are 28% and 19% (Mr. D.S. Martin,
pers. comm.). After
the early studies instigated by Sir John Murray
(Murray, 1904 and 1908; Murray and Pullar, 1910),
few authors published on the fauna inhabiting the
deep sediments of Loch Ness or Loch Morar until
Maitland (1981), Shine and Martin (1988), and Griffiths,
Martin, Shine and Evans (1993). Little work was done on the profundal fauna
of five Scottish lochs during the 1977-80 survey
(Maitland, 1981) but Smith et al. (1981) reviewed what
had been recorded earlier. Dredgings from 90-230 m depths in Loch Ness
by Murray and Pullar collected the bivalve Pisidium, subsequently identified as P. conventus and P. personatum, the lumbriculid Stylodrilus
heringianus, and Chironomus
sp., together with the 'casual' occurrence
of Hydra and
Lymnaea peregra from depths of 120-180 m. Samples taken from c. 200 m depth in Loch Morar contained two lumbriculid
worms, Lumbriculus
variegatus and Stylodrilus
heringianus (Smith et.
al., 1981).
These authors also recorded
that the profundal sediments of Loch Ness, taken
by core, consisted of fine size fractions with clay
(<39 m, 45.8%), fine silt (39-63 m, 18.1%) and sand (63-125 m, 35.4%), low organic content (loss on ignition, 8.9%)
and a pH of 5.4. The paucity of records is largely due to
the difficulties of sampling sediments at the great
depths of these lochs.
As part of the sampling
Vol 105, The Scottish Naturalist: Profundal Fauna of
Loch Ness and Loch Morar p116
programme undertaken by the Loch Ness and Morar Project,
Shine and Martin (1988) designed a variety of collecting
techniques, and compiled a list, not previously
published, of thirty profundal zoobenthic species,
at an average density of 295 individuals/m2, from Loch Ness. Ostracods
formed an important component, comprising over 60%
of the fauna by numbers.
The four species recorded from the deep sediments
(>200 m) of Loch Ness were Candona angulata G.W. Muller, C. candida (O.F. Muller), Cryptocandona reducta (Alm) and Cypria ophthalmica (Jurine) (Griffiths
et al., 1993).
This present paper publishes the full listing of macrobenthic
and meiofaunal species from the profundal sediments
of both lochs, collected over the past ten years
during the Loch Ness and Morar Project's benthic
sampling programme.
Methods
Sampling Sites
The samples from Loch Ness were mostly collected
between June and October of 1982-85, from mid-loch
and in depths between 189 and 230 m, from the areas
Horseshoe Scree to Alltsigh and between Inverfarigaig
to Urquhart Bay (Figure
1a, 7K ). The profundal sites (>200 m) sampled during 1991-93 are plotted
in Figure
2 (9K). Samples from Loch Morar, collected during
July or August in 1978-80 and 1985, were all taken
from depths of 295-310 m in the deep basin between
Meoble Bay and Swordlands (Figure
1b, 7K). Echo-sounders were used to measure the depths
being sampled, as well as to detect and control
the descent of the sampling gear in
situ, especially the Ekman sampler (Figure
3, 16K chart).
Collecting Gear
In the absence of the expensive Jenkins mud-sampler,
several techniques were used to collect samples
from the deep sediments during the earlier sampling
period (1978-85) (Figures 4a,
4b,
4c,and
4d). Unquantitative samples were taken in both lochs,
using a weighted dredge with a fine mesh cod-end
(Figure
4a, 8K) which collected the larger organisms;
additional unquantitative methods, used only in
Loch Ness, were fine-meshed eel-traps on the loch
bottom and a set of gill nets of 50 mm mesh (1.4
m high and 125 m long) kept suspended with the lead-line
resting on the sediments. Left overnight, these caught profundal fish
and some invertebrates. Quantitative samples were mostly taken by a
weighted Ekman grab (15 cm x 15 cm) or, in Loch
Morar only, by a 5.0-litre brass Patalas volume
sampler, which sampled an area 10 cm x 10 cm in
the deep basin of 295-310 m depth. Two special bottom samplers were
designed by Adrian Shine to collect the surface
few centimetres of sediment: a 0.25 m2 quadrat
bottom sampler (Figure
4b, 20K) and a 0.25 m2 scoop bottom sampler (Figure
4c, 17K).
More recent sampling in Loch Ness (1991-93)
was conducted using a new quantitative 10.3 cm diameter
corer (Figure
4d, 7K photo), which permitted sub-samples
to be cut at 1.0 cm intervals, thus giving samples
of 83.3 cm3 for each 1.0 cm depth interval. A second technique used was to dredge the sediment
surface. In
Loch Morar during the 1990s, only the deep basin
(310 m) was sampled, by leaving a buoyed sterilised
hessian sack on the bottom and retrieving it after
two weeks.
Sample Treatment
Profundal samples were kept alive in a refrigerator,
seived, and sorted live as soon as possible; samples
for fine-sorting in London were transported overnight
in a cold container. Some chironomids were kept alive until they
emerged as adults. Only Loch Morar sediment samples were washed
through a series of seives down to 75 micro/m mesh. The most
effective method of quantitative sorting of the
Loch Ness quantitative samples was by visually panning
a series of diluted 10 ml sub-samples until the
whole sample was completed. This was continued until no more live animals
were detected. Sorted animals were preserved and sent to taxonomic specialists
for identification (Tables 1 and 2). The 1990s Loch Ness samples from the corer
were mostly cut into slices, 0.0 - 1.0 and 1.0 -
2.0 cm from the surface mud, and totally sorted. The whole mud sub-sample was examined microscopically,
one spatula load at a time, and diluted to the thinnest
layer possible in a petri dish. The surface dredge samples were seived through
100 m mesh after settlement of larger particles.
The Loch Ness records for the period 1982-85 came from
a total of 37 samples (eight by dredging, six by
the scoop or quadrat samplers, and 23 by Ekman sampler)
and for the 1991-93 period from a total of 11 samples
(seven by dredging and four by coring); the Loch
Morar records for 1978-85 came from a total of 12
samples (eight by Patalas sampler, three by Ekman
sampler, and one by dredging).
Results
Twenty-nine species or genera of profundal
invertebrates, and two species of fish, were recorded
in Loch Ness during the earlier period of sampling,
compared with only seventeen species or genera of
invertebrates in Loch Morar (Table 1). The lesser number of species recorded in
Loch Morar reflects the smaller sampling
Vol 105, The Scottish Naturalist: Profundal Fauna of Loch Ness and Loch
Morar p123
effort, and the fewer collecting techniques
employed, rather than implying any real difference. The species came from 200 m depth. Benthic
collections were made at other depths, but are excluded
from this paper. In Table 1, most of the species collected
were the larger macrobenthic forms present, due
to the methods of collection. That these methods did collect the small
ostracods and copepods probably reflects their numerical
abundance. Some
collecting methods turned out to be selective; for
example, the scoop sampler (Figure
4c) was better at catching cyclopoid copepods,
which swim just above the sediments, than the Ekman
sampler.
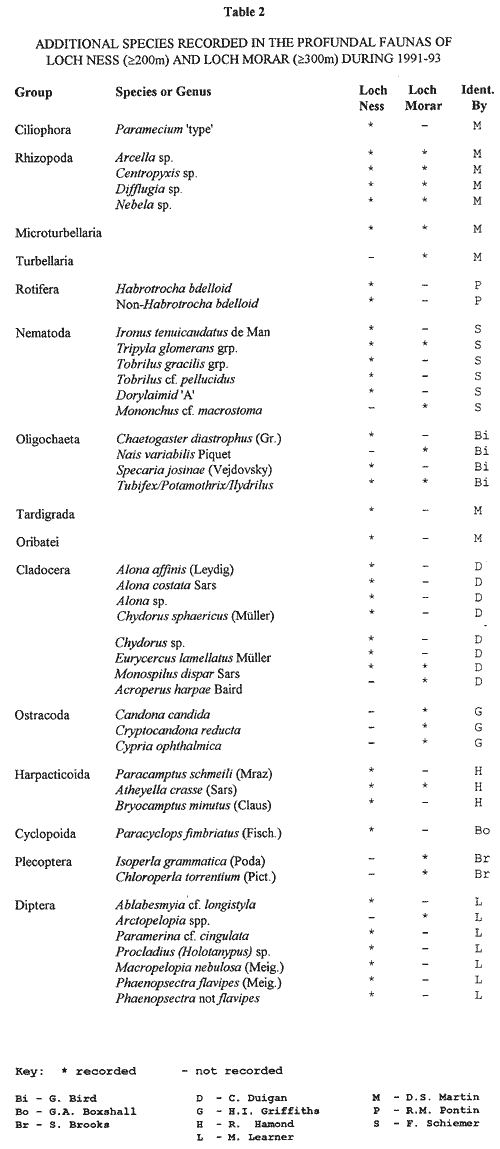
Table 2 lists the species collected during
the more recent sampling series (1991-93), using
better methods of collection (e.g. a corer), with
smaller volumes of mud, more easily sorted accurately
and quantitatively. This list includes additional species/genera
recorded, and also contains more of the smaller
meiobenthic groups of animals which are more difficult
to extract from the mud. This brings the total for Loch Ness to sixty-four
invertebrate species/genera, plus two fish species,
and for Loch Morar to thirty six species/genera. These lists are based on extracting whole
live specimens.
Further species were later added to the list,
by accepting the chironomid head capsules sorted
from the top few cms of Loch Ness core samples. These further species were identified by
Mr. D.P. Gallagher as follows: Heterotrissocladius, Orthocladius, Synorthocladius, Paratrichocladius,
Psectrocladius, Pseudo-chironomus and Tanytarsus.
Comparing
the two lochs, twenty-one species or genera were
recorded in both lochs. With more frequent and more extensive sampling,
it seems likely that the number of species recorded
from Loch Morar will increase. In Loch Lomond, the only other major Scottish
loch in which the profundal benthos has been sampled,
only nine species (four oligochaete, two of Pisidium,
and three chironomid species) were recorded
(Weerekoon, 1956; Slack, 1965; Maitland, 1981).
Discussion
Under the stable environmental conditions
of the profundal (200 m) of Loch Ness, i.e. constant cold (5.6C), darkness, high levels of dissolved oxygen (>80%
saturated), low conductivity (<100 uS.cm-1) and great hydrostatic pressure
(Shine and Martin, 1988), a fauna of surprising
diversity exists, which can be sub-divided as follows:
Vol
105, The Scottish Naturalist: Profundal Fauna
of Loch Ness and Loch Morar p124
(A) The true profundal macrobenthic groups
of chironomids (with 13 species/genera), oligochaetes
(14 species) and the sphaeriid bivalve Pisidium
(four species), each containing a dominant species
within their group.
(B) The true profundal meiobenthic groups
of the ostracods (six species), the nematodes (five
species), harpacticoid copepods (three species)
and cyclopoid copepods (two species), each contributing
dominant species within their group.
(C) A series of macrobenthic
groups, comprising turbellarians (two species),
isopods (two species), amphipods (one species),
and Trichoptera (three species); and meiobenthic
groups, comprising protozoans (five species), microturbellarians
(one species), rotifers (two species), tardigrades
(one species), and oribateids (one species), with
fewer representatives and whose distribution is
not characteristically profundal. The chydorid cladocerans (seven species)
have also been placed in this group, for reasons
given below.
The two fish species, i.e. Lamprey Lampetra sp. and Charr.
Apart from the fish, these organisms are
all living on or in the sediments, or in the water
just above the sediments of very fine fraction (Smith
et al., 1981),
and with an organic content which ranges between
18.29% at Inverfarigaig at 210 m to 33.9% at Invermoriston
at 210 m (Mr. J.D. Hamilton, pers. comm.). The new values are higher than the 8.9% organic
content recorded by Smith et
al. (1981), which came from a single core taken
in the less deep shoulder of Loch Ness between the
North and South Basins where the loch is under the
influence of the River Foyers (Figure
2). In
the profundal, most of the listed species will be
detritivorous, apart from the carnivorous groups
like the turbellarians, cyclopoid copepods, tanypodine
chironomids and fish.
Sub-division
(A)
The chironomids, oligochaetes and species
of Pisidium
belonging to sub-division (A) contribute species
to the profundal macrobenthos in some other deep,
oligotrophic lakes of Europe, e.g. Lake Malaren
(Ahren and Grimas, 1965), Thingvallavatn (Lindegaard,
1992), Lago Maggiore (Lenz, 1954; Della Croce, 1955;
Stella, 1964; Bonomi, 1967; Kuiper, 1981), Lago
di Mergozzo (Bonomi and Ruggin, 1966), and Lake
Geneva (Zschokke, 1911; Juget, 1958; Dussart, 1967);
and of North America, e.g. Great Slave Lake (Rawson,
1953), and the St. Lawrence Great Lakes (Cook and
Johnson, 1974).
Vol 105, The Scottish Naturalist:
Profundal Fauna of Loch Ness and Loch Morar
p125
Sub-division
(B)
Similarly, the sub-division (B) groups contribute
to the profundal meiobenthos in Finnish lakes, e.g.
Paajarvi (Holopainen and Paasivirta, 1977) and Paijanne
(Sarkka, 1992); in Polish lakes (Prejs and Papinska,
1983); and in pre-alpine Austrian lakes, e.g. Vorderer
Finstertaler See (Bretschko, 1973, 1975, 1984),
Piburger See (Pehofer, 1977) and Mondsee (Danielopol
et al., 1988).
Sub-division
(C)
Sub-division (C) is very heterogeneous, and
consists of species able to survive in the profundal
zone but not especially characteristic of it For
example, nine of Fryer's (1985) list of crustacean
species, collected more from large rather than small
water bodies in his survey of 207 lowland Yorkshire
waters, are recorded in Loch Ness or Loch Morar: of these, four are chydorid cladocerans (Alona affinis, Chydorus sphaericus, Eurycercus lamellatus and Acroperus harpae), plus the ostracods Cyclocypris ovum and Cypria ophthalmica, the cyclopoids Paracyclops fimbriatus and Megacyclops viridis, and the isopod Asellus aquaticus. These might be termed vagile (widely dispersive) organisms. Some of sub-division (C) are introduced species: Phagocata woodworthi (turbellarian) (Reynoldson,
Smith and Maitland, 1981) and Crangonyx
pseudogracilis from North America. The presence of Asellus
spp. in the profundal is less surprising when
one considers how the amphipod Pontoporeia affinis dominates the deep
profundal benthos of the St. Lawrence Great Lakes
(Cook and Johnson, 1974) and the Great Slave Lake
(Rawson, 1953) in North America, although to a greater
extent than seems to occur in European lakes. The Trichoptera larvae (and Plecoptera nymphs
in Loch Morar) are probably littoral in origin,
but are able to survive in the well-oxygenated profundal
sediment surface. However, in the few biological studies of
the deep sediments of oligotrophic lakes which include
the profundal meiobenthos, species are recorded
in the profundal which belong to the same animal
groups listed in sub-division (C), plus some others
(Holopainen and Paasivirta, 1977). Many of the sub-division (C) species are
rare or exist as sparse populations able to survive
under the stable environmental conditions of Loch
Ness deep sediments (Giller, 1984).
Sub-division
(D)
The Lamprey was a larva and was caught by the hessian sack technique. More information on the numbers and sizes
of profundal Charr caught can be found in Shine
and Martin (1988) and Shine, Kubecka, Martin and
Duncan (1993).
Vol 105, The Scottish Naturalist:
Profundal Fauna of Loch Ness and Loch Morar
p126
More detailed accounts of the seven groups
in sub-division (A) and sub-division (B), i.e. Chironomidae,
Oligochaeta, Sphaeriidae, Ostracoda, Nematoda, Harpacticoid
Copepoda, and Cyclopoid Copepoda, are as undernoted:
Chironomidae
The taxonomic composition of the profundal Chironomidae
was more diverse in Loch Morar than in Loch Ness,
despite the fewer species collected. The Loch Ness forms belonged to only two
sub-families, the Tanypodinae (six taxa), with predacious
head capsules and capable of feeding on small invertebrates,
and the Chironominae (six taxa), with a non-carnivorous
head capsule, a variety of feeding habits and diets,
and capable of producing haemoglobin and silken
threads for tube-building (Bryce and Hobart, 1972).
Among the Loch Morar taxa, one was tanypodine, three
were chironomine, and two belonged to the Orthocladiinae,
another non-carnivorous sub-family capable of producing
silk but not haemogloblin (that is, without preadaptation
to low oxygen conditions). In Loch Ness the two most numerous species
were the chironomine Sergentia coracina and the tanypodine Procladius spp., and in Loch Morar Sergentia spp. were dominant. The presence of the orthocladine Heterotrissocladius grimshawi and tanytarsine
Stempellinella
spp. in Loch Morar suggests a greater degree of
oligotrophy than in Loch Ness (Cook and Johnson,
1974; Lindegaard, 1992).
Oligochaeta
In both
lochs, three families of oligochaetes were represented:
Naididae (Specaria
josinae - Lake Paijann; Sarkka, 1992), Tubificidae
(Spirosperma ferox and Tubifex
tubifex - Thingvallavatn; Lindgaard, 1992),
and Lumbriculidae (Stylodrilus heringianus and Lumbriculus variegatus - Thingvallavatn;
Lindegaard, 1992), in each case with characteristic
deep-water profundal species (indicated in brackets)
recorded in similar circumstances elsewhere. In the oligotrophic Finnish Lake Paijanne
(Sarkka, 1992), a higher proportion of naidid over
tubificid species indicated a greater degree of
oligotrophy compared with eutrophicated areas of
the lake. The
most abundant oligochaete species was the tubificid
Spirosperma
ferox in Loch Ness, and S.
velutinus in Loch Morar.
Sphaeriidae
According to Odhner (1923) and Kuiper (1974),
Pisidium conventus
is an arctic species which survives in the cold
regions of alpine and deep temperate
Vol 105, The Scottish Naturalist: Profundal Fauna of Loch Ness and Loch
Morar p127
lakes. In Loch Morar it was the only species collected,
and of the four species in Loch Ness, it was the
most abundant species, together with P.
personatum.
Ostracoda
This was the most abundant group collected
in both lochs. Of the four species recorded in Loch
Morar, Cyclocypris ovum and Cypria
ophthalmica are listed by Fryer (1985) as crustacean
species showing a preference for large water bodies.
Candona candida and Cryptocandona reducta, present in both lochs, were less common in
Loch Ness than Cypria
ophthalmica and Candona
angulata (Griffiths et al., 1993), who comment upon the low
ostracod species diversity they found. Danielopol et
al., (1985, 1988) recorded as many as nine species
of ostracods in the profundal of Mondsee, Austria,
two species of which occur in Loch Ness, Candona candida and C. neglecta
(Table 1).
Nematoda
The nematodes listed in Table 2 are the commoner
species collected from the profundal sediments of
Loch Ness 200 m where they attained densities of up to 10,000 individuals/m2 (Mr. D.S. Martin, pers. comm.). It is not an exhaustive list, and more species
(Eumonhystera
filiformis group, Eumonhystera
cf. longicaudatula, Ethmolaimus pratensis group, Aphanolaimus sp. and Dorylaimus
cf. stagnalis)
and greater densities (particularly of Ironus
tenuicaudatus, with densities of up to 20,000
individuals/m2) are found
in lesser depths such as 50-170 m (Dr. F. Schiemer,
pers. comm.). The species listed consist of widely distributed
nematode species known from a wide range of habitats,
such as ponds, rivers, and especially lakes, but
together these species form a nematode association
characteristic of the profundal zone of deep oligotrophic
lakes with fine sediments, low food availability
and good oxygen conditions.
Such nematode associations, with different
species missing depending upon particular local
conditions, have been found in the Austrian alpine
and pre-alpine lakes, such as Vorderer Finstertaler
See (Bretschko, 1984), Attersee (Dudinski, 1979),
and Piburger See (Pehofer, 1977); in Polish alpine
lakes (Prejs, 1977b); in the Finnish oligotrophic
Lake Paajarvi (Holopainen and Paasivirta, 1977);
and in the arctic Lake Char (Prejs, 1977a). It seems that the species composition of
profundal nematodes, and the carrying capacity of
the sediments, are influenced less by interspecific
competition and more by the combined influence of
the nature of the sediments, the degree of oxygenation
and the level of food available (Dr. F. Schiemer,
pers. comm.).
Vol
105, The Scottish Naturalist: Profundal Fauna
of Loch Ness and Loch Morar p128

Vol 105, The Scottish Naturalist:
Profundal Fauna of Loch Ness and Loch Morar
p130
Vol 105, The Scottish Naturalist:
Profundal Fauna of Loch Ness and Loch Morar
p132
Harpacticoid Copepoda
The same four species of harpacticoid copepods
have been found by Holopainen and Paasivirta (1977)
in the deep oligotrophic Lake Paajarvi, along with
profundal nematode and chironomid species associations. In the Finnish Lake Paijanne, Sarkka (1992)
found that Attheyella
crassa showed a marked preference for the well-oxygenated
end of this oligotrophic lake, which is undergoing
eutrophication at the other end.
Cyclopoid Copepoda
Paracyclops
fimbriatus, which
is a crustacean preferring larger water bodies (Fryer,
1985), occurred in both Loch Ness and Loch Morar,
and has been recorded by Dussart (1967) in Lake
Geneva and by Sarkka (1992) in Lake Paijanne, where
the profundal cyclopoids are species of Diacyclops. Another species commonly recorded in
the profundal literature is Megacyclops viridis, which was often taken
in the scoop samples and is a large enough animal
to be a significant predator.
Conclusion
Without further study, it is difficult to
determine whether the profundal benthos of these
two deep Scottish lochs is a random collection of
species populations accumulated over time, or a
structured assemblage of interacting species populations
of all trophic levels inhabiting the deep, cold,
fine-structured and well-oxygenated loch sediments
(Giller, 1984). The profundal benthos is surprisingly species
diverse, and may well represent a saturated equilibrium
community (that is, with filled ecological niches),
as defined by Giller (1984). The age of the lochs, their environmental stability,
their well-oxygenated conditions, and the not too
low levels of organic food available in the sediments,
all support this possibility.
Further biological studies on both the macrobenthos
and meiobenthos should concentrate on quantifying
both the vertical distributions and any differences
between the North and South Basins in Loch Ness. In order to understand the role of the deep
benthos on the loch ecosystem, there is also a need
for life-cycle studies of the dominant species,
by seasonal sampling and experimental rearing, and,
later, unravelling the biotic interactions between
species.
Vol 105, The Scottish Naturalist:
Profundal Fauna of Loch Ness and Loch Morar
p133
Acknowledgements
The Loch Ness and Morar Project would like
to record its deep gratitude to the undernoted experts
who gladly undertook the identification of specimens
sent to them, without which Table 1 and Table 2
would not exist, or who contributed valuable assistance
in other ways:
Dr.
P.C. Barnard (Natural History Museum), Dr. J.A.
Bass (Institute of Freshwater Ecology, I.T.E. Monks
Wood), Dr. G. Bird (National Rivers Authority, Yorkshire
Region), Dr. G.A. Boxshall (Natural History Museum),
Dr. S. Brooks (Natural History Museum), Dr. P.C.
Cranston (Natural History Museum, and C.S.I.R.O.,
Canberra, Australia), Dr. C. Duigan (Countryside
Commission for Wales), Dr. J.P. Ellis (Natural History
Museum), Dr. G. Fryer (Freshwater Biological Association),
Mr. D.P. Gallagher (University of Dublin), Mr. R.B.
Greer (Department of Agriculture and Fisheries for
Scotland), Mr. H.I. Griffiths (University of Leeds),
Mr. J.D. Hamilton (University of Paisley), Dr. R.
Hamond, Dr. P.A. Henderson (Marine Biology Unit,
Fawley), Dr. M.P. Kerney (Natural History Museum),
Dr. M. Ladle (Institute of Freshwater Ecology, Wareham),
Dr. M. Learner (University of Wales), Professor
H. Loffler (University of Vienna), Mr. D.S. Martin
(Loch Ness and Morar Project), Dr. P. Mordan (Natural
History Museum), Dr. L.C.V. Pinder (Institute of
Freshwater Ecology, I.T.E. Monks Wood), Dr. R.M.
Pontin (Royal Holloway University of London), Professor
F. Schiemer (University of Vienna), Dr. R.W. Sims
(Natural History Museum), Dr. I. Strachan (Scottish
Natural Heritage), Mr. A.C. Wheeler (Natural History
Museum), and Dr. J.F. Wright (Institute of Freshwater
Ecology, Wareham).
The
Project also gratefully acknowledges help from Mr.
John Minshull at Loch Ness, and the early enthusiastic
contributions by Mr. Richard Grinvalds to the Loch
Morar species list.
References
Ahren, T. and
Grimas, U. (1965). The
composition of the bottom fauna in two basins of
Lake Malaren. Report,
Institute of Freshwater Research, Drottningholm,
46:
49-57.
Bonomi, G. (1967). L'evoluzione recente del Lago Maggiore rivelata
dalle conspicue modoficazioni del macrobenton profundo. Memorie
della Istituto Italiano di Idrobiologia, 21: 197-212.
Vol 105, The Scottish
Naturalist: Profundal Fauna of Loch Ness and Loch
Morar p134
Bonomi, G. and
Ruggin, D. (1966). Il
macrobenton profundo del Lago di Mergozzo, Italia. Memorie
della Istituto Italiano di Idrobiologia, 20: 153-200.
Bretschko, G. (1973). Benthos
of a high-mountain lake: Nematoda. Proceedings of the International
Association of Theoretical and Applied Limnology,
18:
1421-1428.
Bretschko, G. (1975). Annual benthic biomass
distribution in a high-mountain lake (Vorderer Finstertaler
See, Tyrol, Austria).Proceedings of the International Association of Theoretical and Applied
Limnology, 19: 1279-1285.
Bretschko, G. (1984). Free-living nematodes
of a high-mountain lake (Vorderer Finstertaler See, Tyrol, Austria). I. Monhystera cf.
stagnalis and Ethmolaimus pratensis. Archiv fur Hydrobiologie, 101: 39-72.
Bryce, D. and
Hobart, A.
(1972). The
biology and identification of the larvae of Chironomidae
(Diptera). Entomologist's
Gazette, 23:
175-217.
Cook, D.G. and Johnson, M.G. (1974). Benthic macroinvertebrates of the St. Lawrence
Great Lakes. Journal of the Fisheries Research Board of
Canada, 31: 763-762.
Danielopol, D.L.,
Geiger, W., Tolderer-Farmer, M., Orellana, C.P.
and Terrat, M-N. (1985). The Ostracoda of
Mondsee: spatial and temporal changes during the
last fifty years. In: Contributions
to the Paleolimnology of the Trumer Lakes (Salzburg)
and the Lakes Mondsee, Attersee and Traunsee (Upper
Austria). (Ed. D.L. Danielopol, R. Schmidt and
E. Schultze). Pages
98-121. Mondsee: Eigenverlag Institut fur Limnologie.
Danielopol, D.L.,
Geiger, W., Tolderer-Farmer, M., Orellana, C.P.
and Terrat, M-N. (1988). In search of Cypris and Cythere - a report of the Evolutionary
Ecological Project on limnic Ostracoda from the
Mondsee (Austria). In: Evolutionary Biology of Ostracoda. (Ed. T. Hanai, N. Ikeya and K. Ishizaki). Pages 485-500. Oxford: Elsevier.
Della Croce,
N. (1955). The
conditions of the sedimentation and their relationship
with Oligochaeta populations of Lake Maggiore. Memorie
della Istituto Italiano di Idrobiologia, 8: 39-62.
Dudinski, S. (1979). Das meiobenthos das
Altersee profundals unter besonderdet berucksichtigung
der freilebenden nematoden. Arbeiten Labor Weyregg, 3: 198-208.
Dussart, B.H. (1967). Les Copépodes des Eaux Continentales. Volumes 1 and 2. Paris: N. Boubee et cie.
Fryer, G. (1985). Crustacean
diversity in relation to the size of water bodies:
some facts and problems. Freshwater Biology, 15: 347-361.
Giller, P.S. (1984). Community Structure and the Niche. London: Chapman
& Hall.
Griffiths, H.I.,
Martin, D.S., Shine, A.J. and Evans, J.G. (1993). The ostracod fauna (Crustacea, Ostracoda) of
the profundal benthos of Loch Ness. Hydrobiologia, 254: 111-117.
Vol
105, The Scottish Naturalist: Profundal Fauna
of Loch Ness and Loch Morar p135
Holopainen, I.J.
and Paasivirta, L. (1977). Abundance and biomass of the meiozoobenthos
in the oligotrophic and mesohumic Lake Paajarvi,
southern Finland. Annales
Zoologici Fennici, 14: 124-134.
Juget, J. (1958). Recherche sur la
faune de Fond du Leman et du Lac d'Annecy. Annales
de la Station Centrale de Hydrobiologie Appliquee, 7: 9-96.
Kuiper, J.G.J. (1974). Een pleistocene vondst
van Pisidium conventus Clessin Nederland en de huidige
geografische verspreiding van deze soort in Europa.
Basteria, 38: 27-40.
Kuiper, J.G.J. (1981). Zur frage der geographischen unterarten bei
Pisidien insbesondere bei Pisidium personatum Malm. Archiv
fur Molluskenkunde, 112: 9-19.
Lenz, F. (1954). Die bodenfauna des
Lago Maggiore und ihre lebensbedingungen. Memorie
della Instituto Italiano di Idrobiologia,
8: 273-322.
Lindegaard, C. (1992). Zoobenthos ecology of Thingvallavatn: vertical
distribution, abundance, population dynamics and
production. Oikos,
64: 257-304.
Maitland, P.S. (Ed.) (1981). The Ecology of Scotland's Largest Lochs: Lomond,
Awe, Ness, Morar and Shiel. Monographiae Biologicae, Vol. 44. The Hague: Junk.
Murray, J. (1904). Loch Ness. Part V: Biology of Loch Ness. Geographical Journal, 24: 442-443.
Murray, J. (1908). Bathymetrical survey of the fresh-water lochs
of Scotland Appendix
VII: Notes on the biology of the lochs of the Ness
basin. Geographical
Journal, 31:
64-67.
Murray, J. and
Pullar, L. (1910). Bathymetrical Survey of the Scottish Fresh-Water
lochs. Vols. 1-6. Edinburgh: Challenger Office.
Odhner, N.H. (1923). Mollusca. Pisidium conventus Clessin. Report
of Scientific Results of the Norwegian Expedition
to Noyaya Zemlya 1921. No. 6: 3-7. Kristiania, Norway: Videnskapsselskapet i Kristiania.
Pehofer, H.E. (1977). Bestand und produktion
benthoscher nematoden im Piburger See (Otztal, Tirol). Dissertations of Department of Limnology, University of Innsbruck, 7: 1-87.
Prejs, K. (1977a). The littoral and
profundal benthic nematodes of lakes with different
trophy. Ekologia
Polska,
25: 21-30.
Prejs, K. (1977b). The
species diversity, numbers and biomass of benthic
nematodes in central part of lakes with different
trophy. Ekologia Polska, 25:
31-44.
Prejs, K. and
Papinska, K. (1983). Ecological
characteristics of lakes in north-eastern Poland
versus their trophic gradient. XI. Meiobenthic and near-bottom meiofauna in 20
lakes. Ekologia Polska, 31: 477-493.
Vol 105, The Scottish
Naturalist: Profundal Fauna of Loch Ness and Loch
Morar p136
Rawson, D.S. (1953). The bottom fauna
of Great Slave Lake. Journal of the Fisheries Research Board of
Canada, 10: 486-520.
Reynoldson, T.B.,
Smith, B.D. and Maitland, P.S. (1981). A species of North American triclad (Paludicola;
Turbellaria) new to Britain found in Loch Ness,
Scotland. Journal
of Zoology, 193:
531-539.
Sarkka, J. (1992). Lacustrine profundal
meiobenthos as an environmental indicator. Hydrobiologia, 243/244:
333-340.
Shine, A.J.,
Kubecka, J., Martin, D.S. and Duncan, A. (1993). Fish habitats in Loch Ness. Scottish Naturalist, 105: 237-255.
Shine, A.J. and
Martin, D.S. (1988). Loch Ness habitats observed by sonar and underwater
television. Scottish Naturalist, 100: 111-199.
Slack, H.D. (1965). The profundal fauna
of Loch Lomond, Scotland. Proceedings of the Royal Society of Edinburgh,
Section B, 69: 272-297.
Smith, B.D.,
Cuttle, S.P. and Maitland, P.S. (1981). The profundal zoobenthos. In: The Ecology
of Scotland's Largest Lochs: Lomond, Awe, Ness, Morar and Shiel. (Ed. P.S. Maitland). Monographiae Biologicae, 44: 205-222 The Hague: Junk.
Stella, E. (1964). Il
Megacyclops
viridis Jurine forma batiale de Lago Maggiore. Memorie della Istituto Italiano di Idrobiologia,
17:
57-79.
Weerekoon, A.C.J. (1956). Studies
on the biology of Loch Lomond. 1. The
benthos of Auchentullich Bay. Ceylon Journal of Science, Section B, 7: 1-94.
Zschokke, F. (1911). Die tiefseefauna
der seen Mitteleuropas. Eine geographische faunistische studie. Monographien und Abhandlungen zur Internationalen
Revue der Gesamten Hydrobiologie und Hydrographie,
4: 1-246.
Received
June 1993
Mr. David S. Martin, Loch Ness
and Morar Project,
Loch Ness Centre, DRUMNADROCHIT,
Inverness-shire IV3 6TU.
Mr. Adrian J. Shine, Loch Ness
and Morar Project,
Loch Ness Centre, DRUMNADROCHIT,
Inverness-shire IV3 6TU.
Dr. Annie Duncan, Hydroacoustic
Unit, Department of Biology,
Royal Holloway University of
London, EGHAM, Surrey TW20 0EX.
